Review Article
Small Molecule Modulation of Lipid-Dependent Cellular Processes against Cancer: Fats on the Gunpoint
Aswin Srivatsav T, Manjari Mishra and Shobhna Kapoor*
Department of Chemistry, Indian Institute of Technology Bombay, India
*Corresponding author: Shobhna Kapoor, Department of Chemistry, Indian Institute of Technology Bombay Powai-400076, India
Published: 05 Apr, 2018
Cite this article as: Aswin Srivatsav T, Mishra M, Kapoor
S. Small Molecule Modulation of Lipid-
Dependent Cellular Processes against
Cancer: Fats on the Gunpoint. Clin
Oncol. 2018; 3: 1449.
Abstract
Lewis Thomas in the Lives of a Cell emphasized the complexity encoded by the variety of lipids and their induced structural platforms. Lipid assemblies are complicated biological constituents in that they self assemble non-covalently and create lipid bilayers, within which lipid molecules relocate in a variety of ways. The matrix of cellular membranes is formed by polar lipids, which consist of a hydrophobic and a hydrophilic portion.
Introduction
The propensity of the hydrophobic moieties to self-associate (entropically driven by water)
coupled with the tendency of the hydrophilic moieties to interact with aqueous environments andin
some cases-with each other, forms the physical basis of the spontaneous formation of membranes
[1]. With careful cataloguing of lipids (Lipidomics) we are only beginning to comprehend
the astounding diversity of lipids in the cellular membranes. The membranes in eukaryotic
cells have a wide repertoire of structural lipids. The first one includes glycerophospholipids
such as Phosphatidylcholine (PC), Phosphatidylethanolamine (PE), Phosphatidylserine (PS),
Phosphatidylinositol (PI), and Phosphatidic Acid (PA) [2]. Sphingolipids constitute another class of
structural lipids with their hydrophobic backbone as a ceramide moiety. The major sphingolipids in
mammalian cells are sphingomyelin and glycosphingolipids. Sterols are the major non-polar lipids
of cell membranes representing a unique class, due to their annealed structures that embody a highly
condensed hydrophobic area. Many possible variations in the lipid’s hydrophilic head groups and
hydrophobic tails add a high level of complexity to the existing vast pool of lipids. A considerable
part of our genome (~5 %) continuously synthesizes and regulates this complex array of lipids,
leaving some simple yet profound questions. Why is such a complex diversity of lipids maintained?
Could this imply that cells are continuously trying to create heterogeneity in the membrane? Are
they aiming at phase coexistence; manifested as the existence of domains of coexisting phase(s)?
Many possible variations in the lipid’s hydrophilic head groups and hydrophobic tails add a high
level of complexity to the existing vast pool of lipids. Steric and electrostatic interactions and
hydrophobic mismatch induce phase segregation that forms distinct domains in the bilayer [3-
5]. This ability of lipid membranes to exist in a multitude of phases, characterized by a different
spatial arrangement and motional freedom of each lipid with respect to its neighbors, has lead to
redefining membrane functionality [6]. These phases are influenced by their molecular structure
and environmental conditions like pH, ionic strength, water content, temperature, and pressure;
they have long fascinated scientists in the field of membrane biophysics.
Lipids fulfill many critical requirements in the cell including composing membrane bilayer,
storing energy due to their reduced state, acting as first and second messengers in signal transduction,
providing functional implementations of membrane-proteins structure and function and finally
recognition processes. Advanced Mass-spectroscopy and analytical techniques have allowed
sensitive and highly selective analysis of lipids of diverse chemical structures within complex
biological samples and testified their intimate involvement and their aberrations in many diseases
such as neurodegenerative disorders, infectious diseases and cancer. Interestingly, lipidomics have
revealed cancer type–specific alteration in the lipidome of cells implying critical roles of lipids in
cancer progression and initiation [7-9]. Admitting, most of the drugs in the market against cancer
(also diabetes and inflammation) elicit their effects via binding to their target proteins and regulating
the underlying cancer-related cellular process. However in line with the increasing evidence
elucidating the role of membrane lipids in regulating numerous cellular functions, they have
emerged as attractive molecular targets where in therapies modulating membrane lipids structures
and localization could be developed to control molecular events including changes in cell signaling,
membrane protein function, localization and gene expression related
to various pathological states-so called the “membrane-lipid therapy”
[10,11].
Table 1
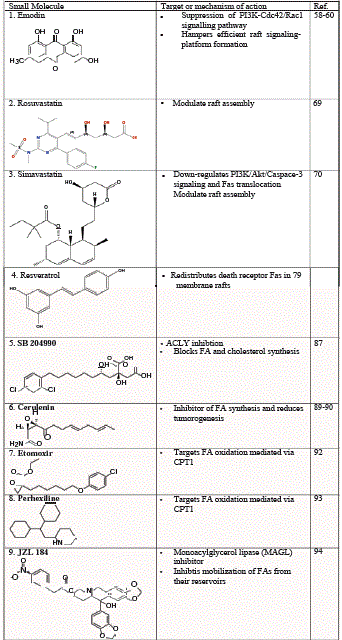
Table 1
Small molecule inhibitors targeting lipid-related cellular pathways for cancer treatment.
Membrane Lipid Micro-Domains as the Cellular Signaling Hot Spots
High compositional complexity modulates interactions and
localizations of lipids in membrane and befittingly influences the
diverse phases lipid can form [12]. The same are the authors of
spatially constrained distinct regions enriched in certain lipids within
the membrane bilayer plane called microdomains [13,14]. One such
microdomains, lipid rafts, are dynamic assemblies of cholesterol and
sphingolipids predominantly in the exoplasmic leaflet of the lipid
bilayer. Underpinning this concept is the propensity, in vitro; of the
saturated hydrocarbon chains intercalating strongly with cholesterol
molecules inducing liquid ordered lo phases [15-17]. The membrane
surrounding lipid rafts is fluid-like due to the presence of unsaturated
phospholipids forming the liquid disordered ld phase. Thus, the raft
domains can be imagined as platforms of lo phases dispersed in the ld
matrix of unsaturated glycerolipids.
These raft-like assemblies are ordered and tightly packed, but are
still fluid due to packing differences. This, in turn, is governed by the
saturation level of the hydrocarbon chains in the rafts compared with
the unsaturated state of fatty acids of phospholipids in the ld phase.
These raft act as membrane scaffolds to house various proteins,
organize receptors and their downstream molecules and hence
regulate a number of membrane-associated signaling pathways
[18,19]. In this regard, post translational modification of proteins such
as addition of GPI anchor, acylation etc. Indispensably modulates the
membrane affinities of proteins including GPI anchored proteins,
epidermal growth factor receptors, and estrogen receptors etc.
Membrane rafts are implicated in regulation of cell differentiation
proliferation, apoptosis, necrosis as well in cancer initiation and
progression [20]. Their involvement in numerous tumor models
such as colon, prostrate, lung and breast has been identified but their
structure, function and associated signaling pathways are still under
intense scrutiny and subject of immense therapeutic interest [21-24].
On the mechanistic level, a cellular signaling event originates
from the raft domains at the membranes with transmission of signals
(chemical, biological and physical in nature) through membrane
bound receptors e.g. Receptor Tyrosine Kinases (RTKs). Stimulation
by the diverse signals promotes receptor dimerization enabling their
phosphorylation mostly via their intrinsic tyrosine kinase activity.
The resulting phosphorylated residues exposed to the cytoplasm
act as docking sites for effect or proteins that harbor specialized
membrane-loving domains—fostering their recruitment to the
membrane. This chain of events induces allosteric changes in the
activity and/or affinities on another module of the recruited molecule
conveying signals deep into the cell and finally leading to regulation
of processes like proliferation, migration, division, and differentiation
among others. Malfunctioning of the cellular signaling both inherent
and induced at any given step in space and time results in unchecked
downstream cellular functions culminating into various diseases,
especially cancer. As an example, we focus on the most fundamental
signaling cascade mediating proliferation and differentiation—
mitogen-activated protein kinase cascade, MAPK signaling, (Figure
1). The pathway gets activated when one of the several extra
cellular ligands (e.g., epidermal growth factors, EGF) bind to the
different plasma membrane-bound receptors, EGFR, leading to the
dimerization and auto-phosphorylation of the cytoplasmic domain
of EGFR (i.e., RTK).
EGFR stimulation promotes binding to adaptor proteins GRB2,
which further binds Son of Sevenless (SOS) [25]. This binding
evokes recruitment of SOS to the plasma membrane, where its close
proximity to the membrane-bound small monomeric G protein,
Ras, forms the prerequisite for Ras activation [26,27]. Ras proteins
are membrane-associated molecular switches that lie at the heart of
MAPK signaling cascade as signaling convergence modules and hold
the place of the first oncogene to be discovered and yet undruggable
to date [28-30].
Activated Ras initiates a series of phosphorylation (and hence
activation) events of protein kinases that act as downstream effect or
proteins of Ras. The most abundant effect or protein of Ras is RAF and
Ras-activated RAF further activates MEK followed by ERK, which
finally travels into the nucleus in its phosphorylated form to activate
transcription factors leading to cell proliferation and differentiation
[31]. This linear description is rather an oversimplified and abridged
depiction of the complex membrane-associated Ras signaling. Misregulation
of Ras signaling e.g. by virtue of failed phosphorylation
events, improper membrane recruitment of effect or proteins,
improper localization with raft domains or mutations account for 50
and 80% of colon and pancreatic cancers, thus emphasizing the
essential role of Ras in normal cell development [32].
Aberrations of Lipids and Lipid Domains in Cancers: Tangible Targets
Pathological, pharmacological, and nutritional situations strongly
regulate lipids in cell with profound biological implications. With
advances in high-throughput lipidomics, precise characterization of
lipid structures is revealing critical lipid alterations in composition
and abundance among various cell types, cancers and surprisingly
during varied cellular processes as well [7,33]. For instance, Eggert
et al demonstrated nice correlation of lipidome changes with cell
cycle, with up to eleven different lipid families (chemically distinct
structures) accumulating in the dividing cells concluding that cells
actively modulate the lipid composition and localization to specific
membrane locations required for a particular cellular event [34].
Cellular lipidome remodeling in cancer is manifold and occurs at
transcriptomic and lipidomic levels with intriguing complexities [35].
Lipid membranes of cancer cells possess relatively higher negative
charge due to increased abundance of PS and PE phospholipids on
the exoplasmic membrane surface [36,37]. On a different note, this
contributes to attenuation of repulsion between polar head groups
leading to denser lipid packing and concomitantly higher rigidity and
poor drug penetration. Thus, exposed PE on the outer membrane of
cancer cell represents a suitable molecular target to develop novel
cancer therapeutics aimed at specific binding to or selective sorting
of PE leading to cancer cell membrane disruption, permeabilization
and finally cell death [38-40]. Cholesterol is another significantly
altered molecule within lipid rafts during cancer, where in the levels
are strikingly increased compared with normal cells [41,42]. Higher
cholesterol leads to a more rigid and hence less permeable cell
membrane [43,44]. In addition to cholesterol, other phospholipid such
as PC, PI etc are also found in increased abundance in cancer cells. The
most foremost effect of elevated cholesterol is higher raft formation
and momentous enrichment of specific proteins and receptors such
as EGFR, IGF-1, CD44, and CD24 etc involved in cellular signaling
mediating tumor progression and invasion [23,45,46]. Thus strategies
involved in modulation of lipid rafts are increasingly becoming
enticing candidates for cancer therapy [47-49]. Down-regulation
of Ceramide metabolism is another strategy found in cancer cells
leading to formation of specialized membrane domains that recruit
specific proteins involved in apoptosis highlighting proteins and
kinases involved in ceramide metabolism as potential cancer targets
[50]. In addition, a wide variety of tumors also show up-regulated
transcripts involved in lipogenesis and cholesterol synthesis pathway,
essential for their development and cancer progression. Lipogenic
enzymes such as acetyl-CoA carboxylase and fatty acid synthase
display a universal increased expression coupled with specific
alterations in lipid messengers (PIs), lipid mediators (leukotrienes)
and structural lipids (GSL) in most tumors [51]. In this review we will
mainly focus on the small molecules that target cancer by modulating
raft assembly, affecting lipid metabolism and regulating lipid sorting
or lipoprotein trafficking inherent in oncogenic signaling (Table 1).
Figure 1
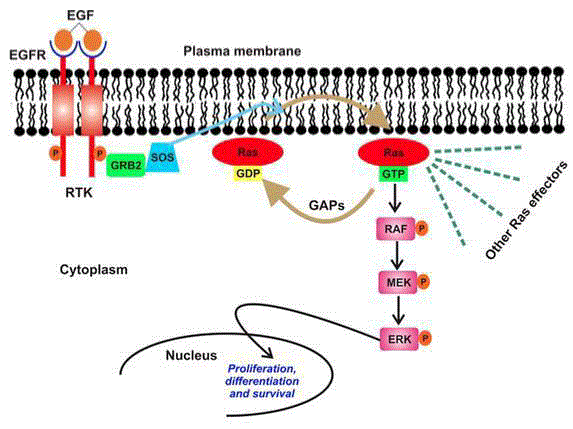
Figure 1
Schematic presentation of MAPK/Ras signaling pathway.
Membrane raft bound-Ras upon activation by variety of extracellular stimuli
undergoes a conformational change facilitated by its membrane localization
in its switch regions, which is then recognized by other downstream effect
or proteins in the pathway. This enables signaling events to get amplified
downstream producing distinct biological outputs raging from cell growth,
differentiation, apoptosis, and vesicle transport. Any kind of alteration in
Ras itself or raft membrane results in various syndromes. Adapted from S.
Kapoor, Dissertation TU Dortmund (2012).
Figure 2
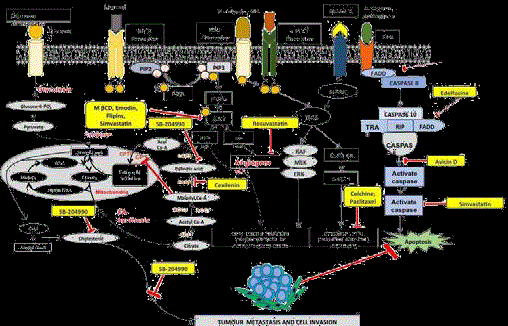
Figure 2
Schematic overview of targeting lipid associated cellular functions in cancer with small molecules.
Small Molecule Chemical Biology Tools
Small molecules targeting specific biomolecules and modulating their structure and activity in vivo has transformed the field of eukaryotic cell biology. Small molecule-mediated inhibition of the function of specific proteins has enabled cell biologists to query their functional roles. Most classic example in this regard is of colchicine and paclitaxel as tubulin depolymerizes and stabilizers respectively, which have provided unprecedented insights into the function of this cytoskeletal protein [52,53]. Development of a toolbox of small molecule inhibitors against cytoskeletal proteins and many more have enabled regulation of their structure, function and localization in such ways that were difficult to achieve solely by genetic approaches. The use of chemical biology tools specifically to study lipid organization offers key advantages. (a) They act fast and their activity can be modulated as a function of dose (b) They may be reversible or not (covalent binders) (c) require no manipulation of the chromosome (d) inhibitors targeting conserved cellular processes may be applicable across a broad range of species. Due to such salient features, they have a great potential in studying the lipid domain organization in live cells, thus permitting insights into the functional role of membrane organization in cancers and other diseases [54,55].
Membrane Raft Modulating Agents in Cancer
Membrane rafts regulate key signaling molecules and proteins implicated in cancer by modulating their association with and localization with lipid membranes including interactions with other membrane-bound proteins [18,20,47,56,57]. Thus small molecule approaches aimed at interrupting the association of such molecules to membrane rafts by interfering with association steps directly or modulating the rafts themselves represents innovative therapeutic ways for prevention and treatment of cancer.
Small Molecules Acting Via Membrane Raft Disruption
Raft disruption using small molecules has been shown to cause decreased cancer cell growth, reduced cellular adhesion and inhibited migration. Among the most promising candidates are flipins, statins, Emodin and Methyl-β-cyclodextrin -MβCD Emodin, 3-methyl-1, 6,8-trihydroxyanthraquinone, inhibits cancer cell migration by mainly suppressing the PI3K-Cdc42/Rac1 signaling pathway [48,58]. Emodin markedly reduces integrin β1 clustering and its colocalization with membrane rafts as judged by cell-based microscopy assays in breast cancer model cell lines [59]. Furthermore, its mechanism of action includes suppression of translocation of integrin β1, and Focal Adhesion Complex (FAC) from cytoplasm to membrane rafts, mainly attributed to reduced cholesterol levels in the membranes, thus hampering efficient raft signaling-platform formation [60]. Thus, the inhibition of membrane raft clustering or activation of raft disruption by Emodin is the underlying mechanism leading to suppression of integrin clustering and FAC formation, and hence halting oncogenic signaling dependent on integrin. Methyl-β-cyclodextrin (MβCD), a cyclic oligosaccharide absorbs cholesterol from the cell membrane and has been shown to impair actin polymerization, cell migration, Akt phosphorylation, protein kinase C translocation and EGF-induced cell adhesion in selective cancer cell models [61,62]. Interestingly, due to the biochemical effects of MβCD, this molecule has been chemically modified to serve as platform for cellular lipid shuffling enabling generation of asymmetric model membrane systems [63-65]. Statins, the first committed inhibitors of the mevalonate pathway act at an early step in the synthesis of cholesterol [66]. The stain family consists of several drugs that are notoriously known for inhibiting oncogenic signaling in cancer cells through disintegration of membrane rafts resulting from reduced cholesterol levels [67-69]. Vastatin inhibits prostate cancer cell growth and inhibits angiogenesis and Simvastatin, another drug belonging to the statin family acts by down-regulating PI3K/Akt/Caspace-3 signaling and Fas translocation mainly by modulation of raft assembly [69,70]. Furthermore, simvastatin blocks Ras-membrane localization, and down modulates H-Ras protein at the post-translational level [71]. It also selectively dissociated latent membrane protein 1 (LMP1) from membrane rafts and reduces activation of NK-κB signaling culminating to apoptosis [72,73]. Surprisingly the small molecules belonging to the statin family, all act predominantly by remodeling membrane rafts composition and organization, however lead to distinct down-stream effects on the oncogenic signaling in various cancer models. This brings to the forefront membrane rafts as “selective cancer therapeutic targets”, with the structure, function and associated raft-signaling pathways being subject of extensive studies. As such, a better understanding of these aspects of raft biology would foster exploitation of membrane rafts for developing personalized cancer therapy for targeting distinct raft-associated oncogenic signaling in various cancers.
Small Molecules Acting by Stabilizing Membrane Domains Involved in Apoptotic Signaling
Recent studies have elucidated membrane rafts to form signaling platforms capable of activating pro- and anti-apoptotic pathways susceptible to pharmacological perturbations aimed at stabilizing these special apoptotic-linked raft domains [74]. Activation of pro-apoptotic pathways begins with activation of pro-apoptotic membrane receptor molecules via oligomerization by agents that promote raft integrity in the absence of receptor ligands. There are two major apoptotic pathways, extrinsic and intrinsic those originate from membrane rafts [75]. The extrinsic pathway is kick-started by death receptors, e.g. Fas. Following stimulation by its ligand–FasL, Fas undergoes clustering and recruits adaptor protein, Fas-associated death domain-containing protein–FADD [76]. FADD interacts with procaspace-8 forming the so-called Death-Inducing Signaling Complex (DISC) that leads to activation of downstream signaling and eventually apoptosis [77]. Activation or clustering of Fas receptor or death receptors in general is critically dependent on membrane rafts to trigger apoptotic signal transduction and is amenable to small-molecule perturbations as follows [78]. Resveratrol is shown to induce apoptosis in colon cancer cells by redistributing Fas among other death rectors in membrane rafts [79]. Avicin D, a plant triterpenoid selectively inhibits growth of tumor cells via activation of caspase pathway, i.e., regulated Fas translocation into membrane rafts and subsequently interactions with FADD and procaspcase-8 to form DISC and hence cause cell apoptosis [80]. Finally along similar lines, Edelfosine (1-O-octadecyl-2-O-methyl-rac-glycero- 3-phosphocholine)–a synthetic lipid, induces apoptotic response by accumulating in the membrane rafts and altering their lipidprotein concentrations and organization [76,81-83]. This leads to co-clustering of FADD and procaspase-8 into membrane rafts and thus activated formation of DISC. Remarkably Edelfosine is highly selective for leukemia cells and solid tumors compared with normal cells, where it targets only the plasma membrane rafts of leukemia cells and endoplasmic rafts of solid tumor cells [84].
Small Molecules Rewiring Lipid Metabolism in Cancer
Cancer cells display a highly distinct metabolic growth profile compared with non-transformed normal cells. The metabolic reprogramming of the enzymes of various pathways of cell growth forms the underlying basis of cancer. One of the most implicated pathways that are heavily tinkered with in cancer is lipid metabolism. Lipid metabolism is linked closely with the glycolytic pathway by virtue of it providing the required starting substrate–acetyl CoA–for Fatty Acid (FA) synthesis. Lipids play key roles in this network, as they are crucial for the formation of cell membranes and also act as signaling messengers. Due to the enormous up-regulated growth rate of cancer cells, relatively larger amounts of lipids are required to keep up with alarming rates of growth, proliferation, energy storage and production of signaling molecules [51,85]. Targeting lipid metabolism encompassing perturbing synthesis, oxidation and mobilization of lipids—is promising strategy in cancer treatment. One of the important steps in lipid metabolism is the formation of fatty acid, which uses acetyl CoA as a substrate. Acetyl CoA is either obtained from the glycolytic pathway via the conversion of pyruvate or by the breakdown of citrate into acetyl CoA and oxaloacetate by cytoplasmic ATP citrate lyase (ACLY). Acetyl CoA binds with malonyl CoA (formed via the carboxylation of acetyl CoA) to form palmitate, which is a starting product of FA synthesis via the enzyme Fatty Acid Synthase (FASN). Inhibitors against ACLY will lead to the reduced production of acetyl CoA and in turn reduce the levels of FAs that are formed. ACLY inhibition has been shown to cause growth suppression and induce apoptosis [86]. SB-204990 is shown to inhibit ACLY and therefore block the synthesis of FA and cholesterol. This causes a block in the cancer cell growth and the suppression of tumor, leading to cell death [87]. The next main step amenable to small molecule targeting is the formation of Palmitate by FASN. Palmitate is then converted by a set of enzymes to form an array of saturated and unsaturated FAs. FASN has been well documented with regard to its role played in cancer and is exploited extensively as anti-cancer target [88]. Targeting FASN has been demonstrated to lead to the de novo FA synthesis in cancer cells to reduce as most normal cells would prefer- the exogenous sources of FA. For example, Cerulenin, an antifungal agent, is one such inhibitor of FA synthesis, which reduces FA synthesis and rescues tumorous cells [89,90]. Another such drug is C75 that has been shown to cause the inhibition of FASN [91].
Small Molecules Targeting Lipid Re-Localization and Lipoprotein Sorting
Next to targeting the enzymes involved in lipid biosynthesis,
targeting lipid oxidation and mobilization/localization are
fruitful therapeutic avenues gaining recent interest. Carnitine
Palmitoyltransferase 1 (CPT1) is an enzyme involved in the β
-oxidation of FAs, where it facilitates the movement of FA-CoA
from the cytosol to the mitochondria across the mitochondrial
membrane. Etomoxir and Perhexiline are two small molecules
shown to be effective against tumors and curb their proliferation
via targeting FA oxidization mediated through CPT1 [92,93]. The
FAs can once successfully translocated can either be diverted for
storage or mobilized from stores as and when needed. The enzymes
involved in these mechanisms have proven to be suitable targets for
cancer therapies. Glycerol-3-Phosphate Acyl Transferases (GPATs)
and its isoforms enable formation of Diacylglycerols (DAG) and
Triacylglycerols (TAG), which are then directed towards storage,
while enzymes like Monoacylglycerol Lipase (MAGL) mobilize
FAs from their reservoirs. CT-30501 inhibits GPATs while JZL184
inhibits MAGL. These small molecules help in suppression of tumor
growth and induce apoptosis respectively [94]. The above are just a
few examples of the use of small molecules to target lipid metabolism
and associated processes in cancer cells and form a firm foundation
of lipid-targeted cancer therapies. One of the salient features of lipid
membranes is the asymmetric distribution of lipids, such lipids PS
and PE. They are largely present in the leaflet facing towards the
cytosol, however upon exposure to the cell surface they act as markers
for signaling pathways. Therefore, this feature of the membrane’s
asymmetric distribution of composing lipids makes it an important
target to fight against much human pathology, and certain lipids are
extensively used as biomarkers against cancer attributed to the fact
of cancer cells expressing high levels of PE and PS exposed on its
outer leaflet. PE acts as a structural component of cell wall as well
as is implicated in many cellular processes like cell division and cell
death, thus a highly sought-after anti cancer target [2]. Cyclotides are
cyclic peptides that have a high affinity to target and bind to PE head
groups modulating their localization and disturbing downstream
cellular functions involving PE. Along with cyclotides, there are two
lantibiotic peptides– Cinnamycin and Duramycin–that are also PE
specific and are produced by gram-positive bacteria. The binding of
both these types of peptides have a membrane disruption effect that
causes cell death [95,96].
Lipid apart from being intimately involved in cellular functions
and cellular signaling as isolated modules, they add another level
of complexity by their covalent attachment to proteins—posttranslational
protein lipidation—that forms the heart of membraneassociated
signaling in cells, e.g. small GTPases, such as Ras, Raf,
and ARFs etc. A classic example of addressing oncogenic signaling
involving lipidated proteins is via targeting the protein lipidation
leading to improper membrane-raft localization of these proteins
causing non-functional signaling platforms and hence subdued
oncogenic signaling. This aspect is greatly exemplified by the class of
lipidated protein-Ras. The lipid moieties attached to the protein consist
of a palmitoyl group and 1-2 farnesyl lipid anchors. As mentioned
above, Ras was the first oncogene to be discovered and is involved in
many human cancers; however small-molecule targeting of Ras still
remains an unmet task in cancer therapy. One of the attempted ways
by which membrane-associated Ras oncogenic signaling has been
targeted is via inhibiting the activity of farnesyl transferase to block
farnesylation of Ras and hence reduce its membrane raft association
and concomitant signaling in cancer cells [97-99]. More than two
decades were invested to exploit this approach as a practical anticancer
therapy but has met with many deadlocks mainly ascribed
to the non-selective nature of farnesyltransferase inhibitors [100].
Amidst such failed attempts, recently Waldmann and co-workers
have demonstrated an exciting alternative to target Ras-associated
cancer by mis-localizing Ras lipoprotein not channelized via blocking
the lipid attachment but by an innovative chemical biology approach
[101]. Lipidated Ras is trafficked through a prenyl-binding protein,
PDEδ in cells that sustains the spatial orientation of the Ras super
family of proteins [102].
Recently high specificity of PDEδ towards K-Ras trafficking to
reach plasma membrane rafts to initiate signaling was demonstrated,
and the same was exploited by designing small molecules such as
Deltarasin and related analogs to block the binding pocket of PDEδ
leading to K-Ras mislocalization and down-regulated cancer signaling
leading to reduces cell proliferation and finally cancer cell death
[101,103-104]. This study provides a proof-of-concept platform and
opens various channels aimed at targeting lipids-mediated cellular
functions in unprecedented ways. Although a lot of work has to be yet
done in identifying cancer specific small molecule against cancer cells,
the effects of presently available small molecule on the membrane
specific organization and signaling are proven as an effective.
Conclusions and Future Directions
The quest for targeting cancer using varied chemical and genetic approaches still is faced with enormous hurdles and generates a unmet need to develop therapeutic approaches inspired by careful inspection of modulated cancer cell attributes. One of the aspects gaining considerable attention recently has been the altered lipid repertoire of cancer cells leading to modulated membrane-dependent cellular processes including membrane organization and cellular signaling, strongly contributing to tumor growth and metastasis and understanding the underlying mechanism behind the same to elucidate potentially novel targets and pathways against cancer. In this review we focused on some of the most promising lipid associated candidates and processes for anti cancer targeting by small molecules. Ranging from targeting of lipid enzymes involved in the lipid metabolic pathway to the proteins and lipids that help in lipid organization, oncogenic lipoprotein sorting and signaling membrane micro domains-rafts have proven to be highly crucial to not only contemplate their therapeutic aspect but also address and unveil specific mechanisms of lipid deregulation in cancer. Further chemical biology investigations on the regulation of membrane lipid-dependent signaling pathways in cancer cell may provide novel targets for therapy and elucidating the role of distinct lipid signaling molecules will offer innovative therapeutic opportunities for development of anti-cancer drugs.
References
- Thomas L. Notes of a biology-watcher. The lives of a cell. N Engl J Med. 1971;284(19):1082.
- Van Meer G, Voelker DR, Feigenson GW. Membrane lipids: where they are and how they behave. Nature reviews Molecular cell biology. 2008;9(2):112-124.
- Roux M, Auzely-Velty R, Djedaini-Pilard F, Perly B. Cyclodextrin induced lipid lateral separation in DMPC membranes: 2H nuclear magnetic resonance study. Biophysical journal. 2002;94(2):715.
- Hartmann W, Galla HJ. Binding of polylysine to charged bilayer membranes. Molecular organization of a lipid· peptide complex. Biochimica et Biophysica Acta (BBA)-Biomembranes. 1978;509(3):474-90.
- Wilkinson DA, Nagle JF. Dilatometric study of binary mixtures of phosphatidylcholines. Biochemistry. 1979;18(19):4244-9.
- Feigenson GW. Phase boundaries and biological membranes. Annu Rev Biophys Biomol Struct. 2007;36:63-77.
- Dória ML, Cotrim CZ, Simões C, Macedo B, Domingues P, Domingues MR, et al. Lipidomic analysis of phospholipids from human mammary epithelial and breast cancer cell lines. Journal of cellular physiology. 2013;228(2):457-68.
- Muir K, Hazim A, He Y, Peyressatre M, Kim DY, Song X, et al. Proteomic and lipidomic signatures of lipid metabolism in NASH-associated hepatocellular carcinoma. Cancer Res. 2013;73(15):4722-31.
- Bosco DA, Fowler DM, Zhang Q, Nieva J, Powers ET, Wentworth Jr P, Lerner RA, Kelly JW. Elevated levels of oxidized cholesterol metabolites in Lewy body disease brains accelerate α-synuclein fibrilization. Nat Chem Biol. 2006;2(5):249-53.
- Escribá PV. Membrane-lipid therapy: a new approach in molecular medicine. Trends Mol Med 2006;12(1):34-43.
- Escribá PV. Membrane-lipid therapy: A historical perspective of membrane-targeted therapies—From lipid bilayer structure to the pathophysiological regulation of cells. Biochim Biophys Acta. 2017;1859(9):1493-506.
- Luzzati V, Tardieu A. Lipid phases: structure and structural transitions. Annu Rev Phys Chem. 1974;25(1):79-94.
- Lingwood D, Simons K. Lipid rafts as a membrane-organizing principle. science. 2010;327(5961):46-50.
- Harder T, Scheiffele P, Verkade P, Simons K. Lipid domain structure of the plasma membrane revealed by patching of membrane components. J Cell Biol. 1998;141(4):929-42.
- Rietveld A, Simons K. The differential miscibility of lipids as the basis for the formation of functional membrane rafts. Biochim Biophys Acta. 1998;1376(3):467-79.
- Nicolini C, Kraineva J, Khurana M, Periasamy N, Funari SS, Winter R. Temperature and pressure effects on structural and conformational properties of POPC/SM/cholesterol model raft mixtures—a FT-IR, SAXS, DSC, PPC and Laurdan fluorescence spectroscopy study. Biochim Biophys Acta. 2006;1758(2):248-58.
- Kapoor S, Werkmüller A, Denter C, Zhai Y, Markgraf J, Weise K, et al. Temperature–pressure phase diagram of a heterogeneous anionic model biomembrane system: Results from a combined calorimetry, spectroscopy and microscopy study. Biochim Biophys Acta. 2011;1808(4):1187-95.
- Simons K, Toomre D. Lipid rafts and signal transduction. Nat Rev Mol Cell Biol. 2000;1(1):31-9.
- Allen JA, Halverson-Tamboli RA, Rasenick MM. Lipid raft microdomains and neurotransmitter signalling. Nat Rev Neurosci. 2007;8(2):128-40.
- Michel V, Bakovic M. Lipid rafts in health and disease. Biol Cell. 2007 Mar 1;99(3):129-40.
- Irwin ME, Bohin N, Boerner JL. Src family kinases mediate epidermal growth factor receptor signaling from lipid rafts in breast cancer cells. Cancer Biol Ther. 2011;12(8):718-26.
- Irwin ME, Mueller KL, Bohin N, Ge Y, Boerner JL. Lipid raft localization of EGFR alters the response of cancer cells to the EGFR tyrosine kinase inhibitor gefitinib. J Cell Physiol. 2011;226(9):2316-28.
- Márquez DC, Chen HW, Curran EM, Welshons WV, Pietras RJ. Estrogen receptors in membrane lipid rafts and signal transduction in breast cancer. Mol Cell Endocrinol. 2006;246(1-2):91-100.
- Palmer CP, Mahen R, Schnell E, Djamgoz MB, Aydar E. Sigma-1 receptors bind cholesterol and remodel lipid rafts in breast cancer cell lines. Cancer Res. 2007;67(23):11166-75.
- Li NA, Batzer A, Daly R, Yajnik V, Skolnik E, Chardin P, et al. Guaninenucleotide-releasing factor hSos1 binds to Grb2 and links receptor tyrosine kinases to Ras signalling. Nature. 1993;363(6424):85-8.
- Kamata T, Feramisco JR. Epidermal growth factor stimulates guanine nucleotide binding activity and phosphorylation of ras oncogene proteins. Nature. 1984;310(5973):147-50.
- Chardin P, Camonis JH, Gale NW, Van Aelst L, Schlessinger J, Wigler MH, et al. Human Sos1: a guanine nucleotide exchange factor for Ras that binds to GRB2. Science. 1993;260(5112):1338-43.
- Vetter IR, Wittinghofer A. The guanine nucleotide-binding switch in three dimensions. Science. 2001;294(5545):1299-304.
- Shih TY, Weeks MO, Young HA, Scolnick EM. Identification of a sarcoma virus-coded phosphoprotein in nonproducer cells transformed by Kirsten or Harvey murine sarcoma virus. Virology. 1979;96(1):64-79.
- Parada LF, Tabin CJ, Shih C, Weinberg RA. Human EJ bladder carcinoma oncogene is homologue of Harvey sarcoma virus ras gene. Nature. 1982;297(5866):474-8.
- Kyriakis JM, App H, Zhang XF, Banerjee P, Brautigan DL, Rapp UR, et al. Raf-1 activates MAP kinase-kinase. Nature. 1992;358(6385):417-21.
- Tidyman WE, Rauen KA. The RASopathies: developmental syndromes of Ras/MAPK pathway dysregulation. A Curr Opin Genet Dev. 2009;19(3):230-6.
- Atilla-Gokcumen GE, Eggert US. A comparative LC-MS based profiling approach to analyze lipid composition in tissue culture systems. Methods Mol Biol. 2014;1232:103-13.
- Atilla-Gokcumen GE, Muro E, Relat-Goberna J, Sasse S, Bedigian A, Coughlin ML, et al. Dividing cells regulate their lipid composition and localization. Cell. 2014;156(3):428-39.
- Muro E, Atilla-Gokcumen GE, Eggert US. Lipids in cell biology: how can we understand them better?. Mol Biol Cell. 2014;25(12):1819-936.
- Emoto K, Kobayashi T, Yamaji A, Aizawa H, Yahara I, Inoue K, et al. Redistribution of phosphatidylethanolamine at the cleavage furrow of dividing cells during cytokinesis. Proc Natl Acad Sci. 1996;93(23):12867-72.
- Emoto K, Toyama-Sorimachi N, Karasuyama H, Inoue K, Umeda M. Exposure of phosphatidylethanolamine on the surface of apoptotic cells. Exp Cell Res. 1997;232(2):430-4.
- Kenis H, Reutelingsperger C. Targeting phosphatidylserine in anti-cancer therapy. Curr Pharm Des 2009;15(23):2719-23.
- Chidley C, Trauger SA, Birsoy K, O'Shea EK. The anticancer natural product ophiobolin A induces cytotoxicity by covalent modification of phosphatidylethanolamine. Elife. 2016;5:e14601.
- Wang CK, Wacklin HP, Craik DJ. Cyclotides insert into lipid bilayers to form membrane pores and destabilize the membrane through hydrophobic and phosphoethanolamine-specific interactions. J Biol Chem. 2012;287(52):43884-98.
- Toppozini L, Meinhardt S, Armstrong CL, Yamani Z, Kučerka N, Schmid F, et al. Structure of cholesterol in lipid rafts. Phys Rev Lett. 2014;113(22):228101.
- Zalba S, ten Hagen TL. Cell membrane modulation as adjuvant in cancer therapy. Cancer Treat Rev. 2017;52:48-57.
- Krivanek R, Okoro L, Winter R. Effect of cholesterol and ergosterol on the compressibility and volume fluctuations of phospholipid-sterol bilayers in the critical point region: a molecular acoustic and calorimetric study. Biophys J. 2008;94(9):3538-48.
- Bernsdorff C, Wolf A, Winter R, Gratton E. Effect of hydrostatic pressure on water penetration and rotational dynamics in phospholipid-cholesterol bilayers. Biophys J. 1997;72(3):1264-77.
- Simons K, Ikonen E. Functional rafts in cell membranes. Nature. 1997;387(6633):569-72.
- Zhuang L, Lin J, Lu ML, Solomon KR, Freeman MR. Cholesterol-rich lipid rafts mediate akt-regulated survival in prostate cancer cells. Cancer Res. 2002;62(8):2227-31.
- Mollinedo F, Gajate C. Lipid rafts as major platforms for signaling regulation in cancer. Adv Biol Regul. 2015;57:130-46.
- Hryniewicz-Jankowska A, Augoff K, Biernatowska A, Podkalicka J, Sikorski AF. Membrane rafts as a novel target in cancer therapy. Biochim Biophys Acta. 2014;1845(2):155-65.
- Yang YF, Jan YH, Liu YP, Yang CJ, Su CY, Chang YC, et al. Squalene synthase induces tumor necrosis factor receptor 1 enrichment in lipid rafts to promote lung cancer metastasis. Am J Respir Crit Care Med. 2014;190(6):675-87.
- Yandım MK, Apohan E, Baran Y. Therapeutic potential of targeting ceramide/glucosylceramide pathway in cancer. Cancer Chemother Pharmacol. 2013;71(1):13-20.
- Beloribi-Djefaflia S, Vasseur S, Guillaumond F. Lipid metabolic reprogramming in cancer cells. Oncogenesis. 2016;5(1):e189.
- Peterson JR, Mitchison TJ. Small molecules, big impact: a history of chemical inhibitors and the cytoskeleton. Chem Biol. 2002;9(12):1275-85.
- Ward GE, Carey KL, Westwood NJ. Using small molecules to study big questions in cellular microbiology. Cell Microbiol. 2002;4(8):471-82.
- Fassy F, Dureuil C, Lamberton A, Mathieu M, Michot N, Ronan B, et al. In vitro characterization of Vps34 lipid kinase inhibition by small molecules. Methods Enzymol. 2017;587:447-64.
- Lum KM, Sato Y, Beyer BA, Plaisted WC, Anglin JL, Lairson LL, et al. Mapping Protein Targets of Bioactive Small Molecules Using Lipid-Based Chemical Proteomics. ACS Chem Biol. 2017;12(10):2671-81.
- Kapoor S, Weise K, Erlkamp M, Triola G, Waldmann H, Winter R. The role of G-domain orientation and nucleotide state on the Ras isoformspecific membrane interaction. Eur Biophys J. 2012;41(10):801-13.
- Weise K, Kapoor S, Denter C, Nikolaus J, Opitz N, Koch S, et al. Membrane-mediated induction and sorting of K-Ras microdomain signaling platforms. J Am Chem Soc. 2011;133(4): 880-7.
- Huang Q, Shen HM, Ong CN. Emodin inhibits tumor cell migration through suppression of the phosphatidylinositol 3-kinase-Cdc42/Rac1 pathway. Cell Mol Life Sci. 2005;62(10):1167-75.
- Huang Q, Shen HM, Shui G, Wenk MR, Ong CN. Emodin inhibits tumor cell adhesion through disruption of the membrane lipid Raft-associated integrin signaling pathway. Cancer Res. 2006;66(11):5807-15.
- Huang Q, Shen HM, Ong CN. Inhibitory effect of emodin on tumor invasion through suppression of activator protein-1 and nuclear factorκB. Biochem Pharmacol. 2004;68(2):361-71.
- Liu Y, Sun R, Wan W, Wang J, Oppenheim JJ, Chen L, et al. The involvement of lipid rafts in epidermal growth factor-induced chemotaxis of breast cancer cells. Mol Membr Biol. 2007;24(2):91-101.
- Wu Y, Rizzo V, Liu Y, Sainz IM, Schmuckler NG, Colman RW. Kininostatin associates with membrane rafts and inhibits αvβ3 integrin activation in human umbilical vein endothelial cells. Arterioscler Thromb Vasc Biol. 2007;27(9):1968-75.
- Chiantia S, Schwille P, Klymchenko AS, London E. Asymmetric GUVs prepared by MβCD-mediated lipid exchange: an FCS study. Biophys J. 2011;100(1):L1-3.
- Lin Q, London E. Preparation of artificial plasma membrane mimicking vesicles with lipid asymmetry. PLoS One. 2014 Jan 28;9(1):e87903.
- Crowley JT, Toledo AM, LaRocca TJ, Coleman JL, London E, Benach JL. Lipid exchange between Borrelia burgdorferi and host cells. PLoS Pathog. 2013;9(1):e1003109.
- Roy M, Kung HJ, Ghosh PM. Statins and prostate cancer: role of cholesterol inhibition vs. prevention of small GTP-binding proteins. Am J Cancer Res. 2011;1(4):542-61.
- Chan KK, Oza AM, Siu LL. The statins as anticancer agents. Clin Cancer Res. 2003;9(1):10-9.
- Osmak M. Statins and cancer: current and future prospects. Cancer Lett. 2012;324(1):1-12.
- Wang C, Tao W, Wang Y, Bikow J, Lu B, Keating A, et al. Rosuvastatin, identified from a zebrafish chemical genetic screen for antiangiogenic compounds, suppresses the growth of prostate cancer. Eur Urol. 2010;58(3):418-26.
- Wu H, Jiang H, Lu D, Xiong Y, Qu C, Zhou D, et al. Effect of simvastatin on glioma cell proliferation, migration, and apoptosis. Neurosurgery. 2009;65(6):1087-97.
- Khanzada UK, Pardo OE, Meier C, Downward J, Seckl MA, Arcaro A. Potent inhibition of small-cell lung cancer cell growth by simvastatin reveals selective functions of Ras isoforms in growth factor signalling. Oncogene. 2006;25(6):877-87.
- Mosialos G, Birkenbacht M, Yalamanchill R, Van Arsdale T, Ware C, Kleff E. The Epstein-Barr virus transforming protein LMP1 engages signaling proteins for the tumor necrosis factor receptor family. Cell. 1995;80(3):389-99.
- Katano H, Pesnicak L, Cohen JI. Simvastatin induces apoptosis of Epstein-Barr virus (EBV)-transformed lymphoblastoid cell lines and delays development of EBV lymphomas. Proc Natl Acad Sci U S A. 2004;101(14):4960-5.
- George KS, Wu S. Lipid raft: A floating island of death or survival. Toxicol Appl Pharmacol. 2012;259(3):311-9.
- Gajate C, Gonzalez-Camacho F, Mollinedo F. Lipid raft connection between extrinsic and intrinsic apoptotic pathways. Biochem Biophys Res Commun. 2009;380(4):780-4.
- Gajate C, Gonzalez-Camacho F, Mollinedo F. Involvement of raft aggregates enriched in Fas/CD95 death-inducing signaling complex in the antileukemic action of edelfosine in Jurkat cells. PLoS One. 2009;4(4):e5044.
- Peter ME, Krammer PH. The CD95 (APO-1/Fas) DISC and beyond. Cell Death Differ. 2003;10(1):26-35.
- Gajate C, Mollinedo F. Cytoskeleton-mediated death receptor and ligand concentration in lipid rafts forms apoptosis-promoting clusters in cancer chemotherapy. J Biol Chem. 2005;280(12):11641-7.
- Delmas D, Rebe C, Micheau O, Athias A, Gambert P, Grazide S, et al. Redistribution of CD95, DR4 and DR5 in rafts accounts for the synergistic toxicity of resveratrol and death receptor ligands in colon carcinoma cells. Oncogene. 2004;23(55):8979-86.
- Xu ZX, Ding T, Haridas V, Connolly F, Gutterman JU. Avicin D, a plant triterpenoid, induces cell apoptosis by recruitment of Fas and downstream signaling molecules into lipid rafts. PLoS One. 2009;4(12):e8532.
- Gajate C, Mollinedo F. Lipid rafts, endoplasmic reticulum and mitochondria in the antitumor action of the alkylphospholipid analog edelfosine. Anticancer Agents Med Chem. 2014;14(4):509-27.
- Ausili A, Torrecillas A, Aranda FJ, Mollinedo F, Gajate C, CorbalánGarcía S, et al. Edelfosine is incorporated into rafts and alters their organization. J Phys Chem B. 2008;112(37):11643-54.
- Gajate C, Mollinedo F. Edelfosine and perifosine induce selective apoptosis in multiple myeloma by recruitment of death receptors and downstream signaling molecules into lipid rafts. Blood. 2007;109(2):711-9.
- Gajate C, Matos-da-Silva M, Dakir EL, Fonteriz RI, Alvarez J, Mollinedo F. Antitumor alkyl-lysophospholipid analog edelfosine induces apoptosis in pancreatic cancer by targeting endoplasmic reticulum. Oncogene. 2012;31(21):2627-39.
- Röhrig F, Schulze A. The multifaceted roles of fatty acid synthesis in cancer. Nat Rev Cancer. 2016;16(11):732-49.
- Migita T, Okabe S, Ikeda K, Igarashi S, Sugawara S, Tomida A, et al. Inhibition of ATP citrate lyase induces triglyceride accumulation with altered fatty acid composition in cancer cells. Int J Cancer. 2014;135(1):37- 47.
- Pearce NJ, Yates JW, Berkhout TA, Jackson B, Tew D, Boyd H, et al. The role of ATP citrate-lyase in the metabolic regulation of plasma lipids. Hypolipidaemic effects of SB-204990, a lactone prodrug of the potent ATP citrate-lyase inhibitor SB-201076. Biochem J. 1998;334:113–9.
- Menendez JA, Lupu R. Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nat Rev Cancer. 2007;7(10):763-77.
- Shiragami R, Murata S, Kosugi C, Tezuka T, Yamazaki M, Hirano A, et al. Enhanced antitumor activity of cerulenin combined with oxaliplatin in human colon cancer cells. Int J Oncol. 2013;43(2):431-8.
- Zheng, B.; Zhu, S.; Wu, X. ACS Chem Biol 2015, 10, 115.
- Kuhajda FP, Pizer ES, Li JN, Mani NS, Frehywot GL, Townsend CA. Synthesis and antitumor activity of an inhibitor of fatty acid synthase. Proc Natl Acad Sci U S A. 2000;97(7):3450-4.
- Pike LS, Smift AL, Croteau NJ, Ferrick DA, Wu M. Inhibition of fatty acid oxidation by etomoxir impairs NADPH production and increases reactive oxygen species resulting in ATP depletion and cell death in human glioblastoma cells. Biochim Biophys Acta. 2011;1807(6):726-34.
- Estañ MC, Calviño E, Calvo S, Guillén-Guío B, del Carmen BoyanoAdánez M, de Blas E, et al. Apoptotic efficacy of etomoxir in human acute myeloid leukemia cells. Cooperation with arsenic trioxide and glycolytic inhibitors, and regulation by oxidative stress and protein kinase activities. PloS one. 2014;9(12):e115250.
- Lysenko LV, Kim J, Henry C, Tyrtyshnaia A, Kohnz RA, Madamba F, et al. Monoacylglycerol lipase inhibitor JZL184 improves behavior and neural properties in Ts65Dn mice, a model of down syndrome. PloS one. 2014;9(12):e114521.
- Märki F, Hänni E, Fredenhagen A, van Oostrum J. Mode of action of the lanthionine-containing peptide antibiotics duramycin, duramycin B and C, and cinnamycin as indirect inhibitors of phospholipase A2. Biochem Pharmacol. 1991;42(10):2027-35.
- Svangård E, Burman R, Gunasekera S, Lövborg H, Gullbo J, Göransson U. Mechanism of action of cytotoxic cyclotides: cycloviolacin O2 disrupts lipid membranes. J Nat Prod. 2007;70(4):643-7.
- Agrawal AG, Somani RR. Farnesyltransferase inhibitor as anticancer agent. Mini Rev Med Chem. 2009;9(6):638-52.
- Frassanito MA, Mastromauro L, Cusmai A, Dammacco F. Blockade of the Ras pathway by manumycin, a farnesyltransferase inhibitor, overcomes the resistance of myeloma plasma cells to Fas-induced apoptosis. Clin Exp Med. 2005;4(4):174-82.
- Kohl NE, Mosser SD, Giuliani EA, Pompliano DL, Graham SL, Smith RL, et al. Selective inhibition of ras-dependent transformation by a farnesyltransferase inhibitor. Science. 1993;260(5116):1934-7.
- Ding H, McDonald JS, Yun S, Schneider PA, Peterson KL, Flatten KS, et al. Farnesyltransferase inhibitor tipifarnib inhibits Rheb prenylation and stabilizes Bax in acute myelogenous leukemia cells. haematologica. 2014;99(1):60-9
- Zimmermann G, Papke B, Ismail S, Vartak N, Chandra A, Hoffmann M, et al. Small molecule inhibition of the KRAS–PDEδ interaction impairs oncogenic KRAS signalling. Nature. 2013;497(7451):638-42.
- Schmick M, Kraemer A, Bastiaens PI. Ras moves to stay in place. Trends Cell Biol. 2015;25(4):190-7.
- Chandra A, Grecco HE, Pisupati V, Perera D, Cassidy L, Skoulidis F, et al. The GDI-like solubilizing factor PDEδ sustains the spatial organization and signalling of Ras family proteins. Nat Cell Biol 2012;14(2):148-58.
- Schmick M, Vartak N, Papke B, Kovacevic M, Truxius DC, Rossmannek L, et al. KRas localizes to the plasma membrane by spatial cycles of solubilization, trapping and vesicular transport. Cell. 2014;157(2):459-71.
- Martín‐Gago P, Fansa EK, Klein CH, Murarka S, Janning P, Schürmann M, et al. A PDE6δ‐KRas Inhibitor Chemotype with up to Seven H‐Bonds and Picomolar Affinity that Prevents Efficient Inhibitor Release by Arl2. Angew Chem. 2017;129(9):2463-8.